
Non-Human Primates (NHP)
Non-human primates (NHPs) are used as models in biomedical research because of their close genetic, physiological, and neurological similarities to humans. This makes them especially valuable for studying complex biological processes, such as brain function, immune responses, and infectious diseases, that cannot be accurately replicated in more distant species like rodents.
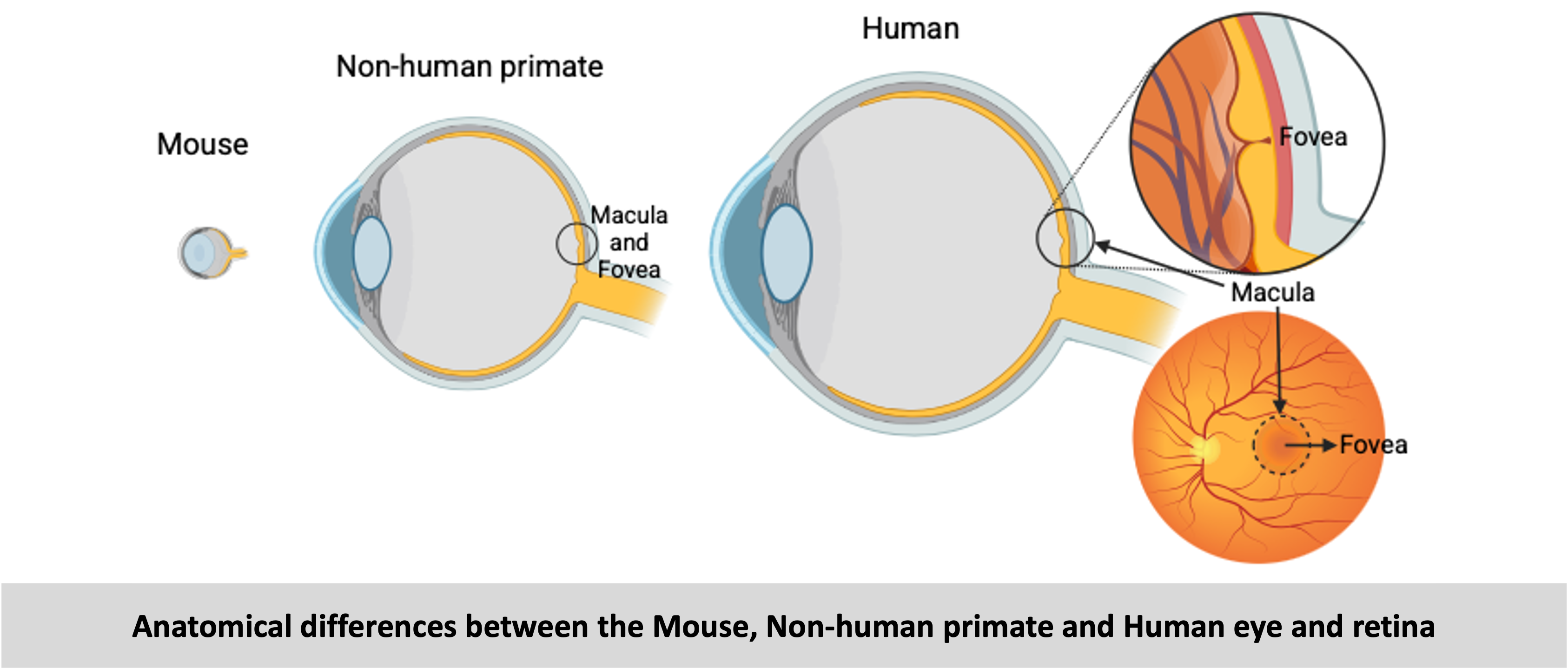
There are many rodent models (mice and rats) of retinal degeneration (RD) that serve well to study retinal diseases and develop some therapies. However, there are several anatomical differences between the rodent and human retina such as the size of the eye, the size and shape of the lens, the relative amount of vitreous and most importantly the presence of a central cone-rich region called macula and a cone-only region called fovea. The macula and fovea are responsible for high-acuity vision and these regions are absent in the rodents which affect how disease features appear and progress in these models, thereby making rodents unsuitable for testing translational therapies.
The non-human primate (NHP) retina is closest to humans and best suited for testing therapies before clinical trials. However, there is a lack of relevant NHP degeneration models that can be used for testing therapies. We addressed this unmet need by using different bioengineering strategies to develop three distinct inducible NHP models of RD –
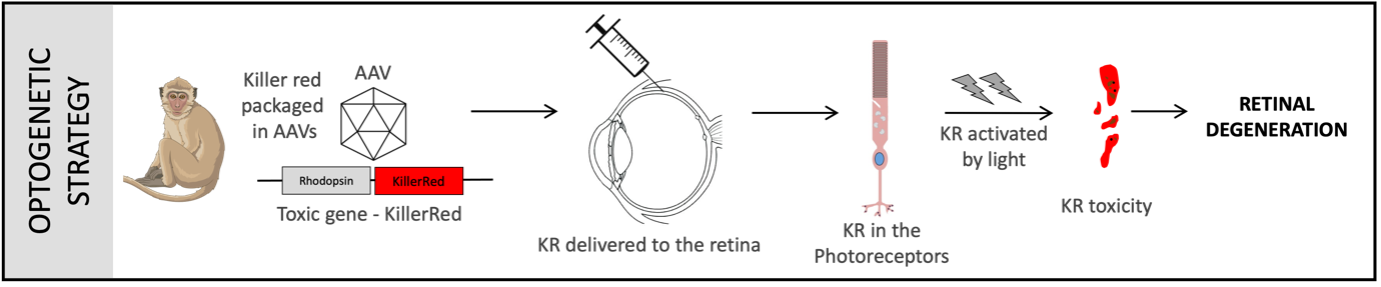
Optogenetic model: In this model an optogene (a gene that can be activated by light) is packaged in AAVs and delivered to the NHP retina by subretinal injections. The optogene is specifically expressed in the rod photoreceptors due to the presence of a rod cell-specific promoter in the transgene design. The optogene can be activated by using light of a specific wavelength (green light of 565nm). The transparency of the eye allowing light to reach the retinal tissue enables optogene activation by exposing the eye to a light source of 565nm. Upon light activation the optogene becomes toxic and results in photoreceptor-cell death, resulting in cell loss and eventually retinal degeneration.
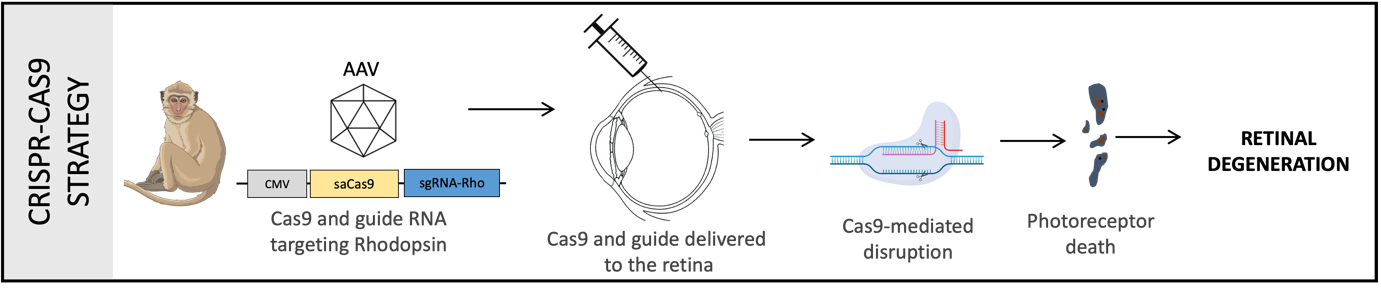
Crispr-Cas9 model: This strategy involves the use of the Crispr-Cas9 system to disrupt the Rhodopsin gene. A Crispr guide is designed against the Rhodopsin gene, packaged into AAV vectors and delivered to the NHP retina by subretinal injections. Rhodopsin is a light-sensitive receptor protein found in the rod photoreceptor cells of the retina and plays a key role in visual phototransduction, the process by which light signal is detected and converted into electrical signal. Mutation in the Rhodopsin gene results in absence of this protein, inability to carry out phototransduction, loss of photoreceptor cells and eventual retinal degeneration. Mutation in the Rhodopsin gene is the most frequent mutation occurring in patients with the disease Retinitis Pigmentosa (RP), and hence this model mimics the RP disease features and progression.
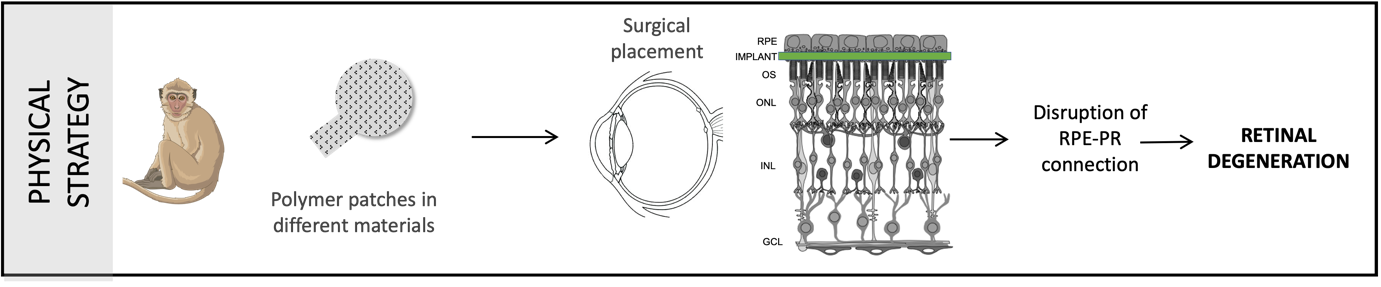
Physical model: This strategy involves using a polymer patch to create a physical separation between the photoreceptors and Retinal Pigment Epithelium (RPE). A polymer patch created with the same materials used in retinal prosthesis/ implants was surgically placed in the subretinal space of the macular region (close to the fovea but without including it). The photoreceptors depend on the RPE for their general maintenance and function. So, when this connection is disrupted, photoreceptors start dying eventually causing retinal degeneration.
The models we have created can be used for testing a wide range of these therapeutic strategies. Degeneration of the retina occurs in stages and the chosen vision restoration method largely depends on the type and stage of the disease. In the earlier stages, when the retinal cells are still present neuroprotective factors can be provided to slow down the progression of the disease and gene replacement or editing can be used to compensate for the effect of gene mutations. On the other hand, in later stages when the retinal cells are mostly lost, mutation-independent vision restoration therapies such as stem cell therapy, optogenetic therapy and retinal prosthesis/ implants could help in restoring some useful vision. Additionally, specific therapies can control symptoms that arise from perturbed cellular mechanisms such as oxidative stress, inflammation or neovascularization.
Evaluation of immune responses to therapy: Injection of viruses, transgenes and other therapeutic agents can trigger an immune response which can reduce the efficacy of the treatment or result in inflammation compromising the safety of the treatment. We used the NHP models that I generated and other NHPs that received ocular gene therapies to study such immune responses.
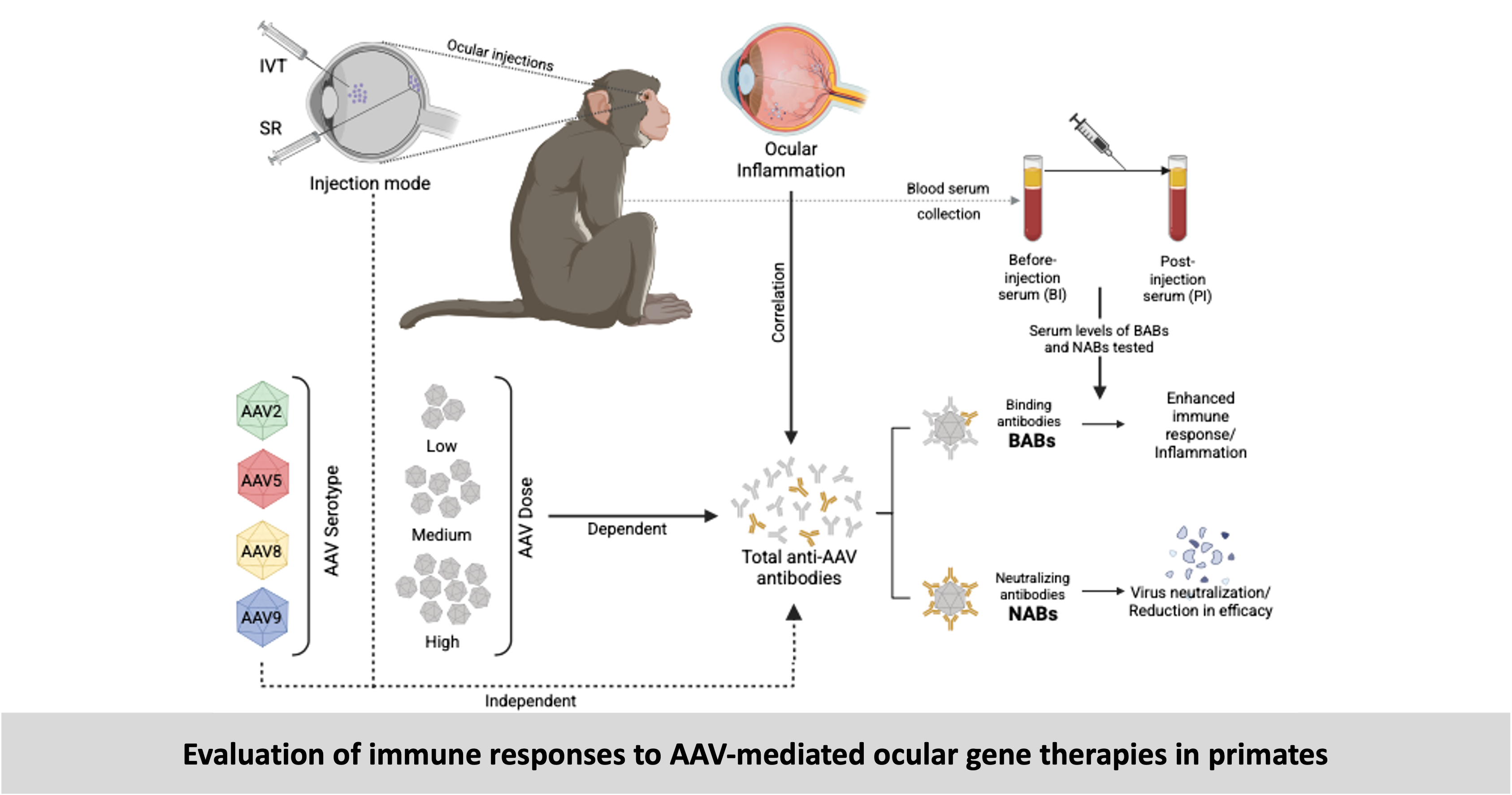
-
Read the details of my projects using NHP models -
- Inducible non-human primate models of retinal degeneration for testing end stage therapies and understanding disease mechanisms
Ail D, Nava D, Hwang I, Brazhnikova E, Nouvel-Jaillard C, Dentel A, Joffris C, Rousseau L, Dégardin J, Bertin S, Sahel JA, Goureau O, Picaud S, Dalkara D
Science Advances, 2023. Access article here - Systemic and local immune response to intraocular AAV vector administration in non-human primates
Ail D, Ren D, Brazhnikova E, Nouvel-Jaillard C, Bertin S, Fisson S, Dalkara D
Molecular Therapy – Methods and Clinical Development, 2022. Access article here - Pre-existing neutralizing antibodies against different adeno-associated virus serotypes in large animal models
Ail D and Dalkara D
Advances in Experimental Medicine and Biology, 2023. Access article here
Rodent models (Mouse and Rats)
Mice are widely used as models in biomedical research because they share a high degree of genetic similarity with humans, are small and inexpensive to maintain, and reproduce quickly, allowing for large-scale studies across generations. Their genomes are well-characterized and can be easily manipulated using genetic engineering techniques to study the roles of specific genes in health and disease. Despite some biological differences from humans, their versatility and the wealth of available research tools make them one of the most important model organisms in medical science. I have developed and worked with a variety of mouse models for over a decade and I have also had the need to perform experiments with rats for some of my projects.
The mouse is a widely used model for studying the retina and retinal degeneration due to its well-characterized genetics, anatomical similarity to the human retina, and the availability of numerous transgenic and knockout strains. Many genes implicated in human retinal diseases, such as retinitis pigmentosa and age-related macular degeneration, have homologs in mice, allowing us to replicate and investigate disease mechanisms in vivo. Mice also offer the advantage of advanced imaging techniques and functional assays, such as fundus imaging, optical coherence tomography (OCT) and electroretinography (ERG), to monitor retinal structure and function over time. These models have been instrumental in understanding the molecular pathways of retinal degeneration and in developing gene therapies and other potential treatments.
Genetic models: For my work on molecular mechanisms of neurodegeneration and neuroprotection, I have relied on the several well-characterized genetic mouse models of retinal degeneration such as rd1 and rd10 mouse models that enable physiological and molecular studies of retinal degeneration. Compared to the rd1 model, rd10 mice have a slower rate of degeneration making them more suitable for therapeutic intervention studies, including gene therapy and neuroprotection. The rd10 model has been instrumental in advancing our understanding of photoreceptor degeneration mechanisms and testing potential treatments for inherited retinal diseases.
Conditional models: We have generated conditional knockdown in mice using the Cre-loxP system. This strategy allows precise control over where and when a gene is knocked down, making it a powerful tool for studying gene function in development, physiology, and disease. Our goal was to test the effect of hypoxia on the photoreceptors by stabilizing the transcription fcators called Hypoxia Inducible Factors (HIFs) in the photoreceptors. In a mouse model (Vhlflox) LoxP sites were inserted flanking (or floxing) the Vhl gene. Another mouse model (Opsin-Cre) expressed the Cre recombinase under the control of a rod photorecptor-sepcific promoter called Opsin. The breeding of these two models resuled in the conditional knock-down of Vhl gene in the rod pgotorecptors which lead to the stabilization of HIFs in these cells. These models helped in better understanding of the role of HIFs in rerinal degeneration.
Inducible models: In addition to genetic models, I have used and developed inducible mouse models to study different mechanisms or test therapies. For the development of the inducible NHP models, the intial proof-of-concept models were developed in rodents resulting in an optogenetically inducible mouse model, a Crispr-Cas9 mouse model and a physical patch-mediated rat model.
Another inducible model that I have extensively used is the light damage mouse model of retinal degeneration wherein photoreceptor cell death is induced by exposure to intense or prolonged light, mimicking aspects of human retinal diseases. In this model, mice—typically albino or light-sensitive strains—are exposed to high-intensity light (often >10,000 lux) for a defined period, leading to oxidative stress, inflammation, and apoptosis, primarily in rod photoreceptors.
To study acute hypoxia (short-term) hypoxia, mice were exposed to hypoxic conditions, in contrast to the study of chronic and long-term hypoxia for which we developed the conditional knock-down models. Exposing mice to hypoxic conditions allow us to assess the effects of low oxygen conditions on physiological responses such as changes in vascularization, oxidative stress, gene expression and inflammatory pathways.
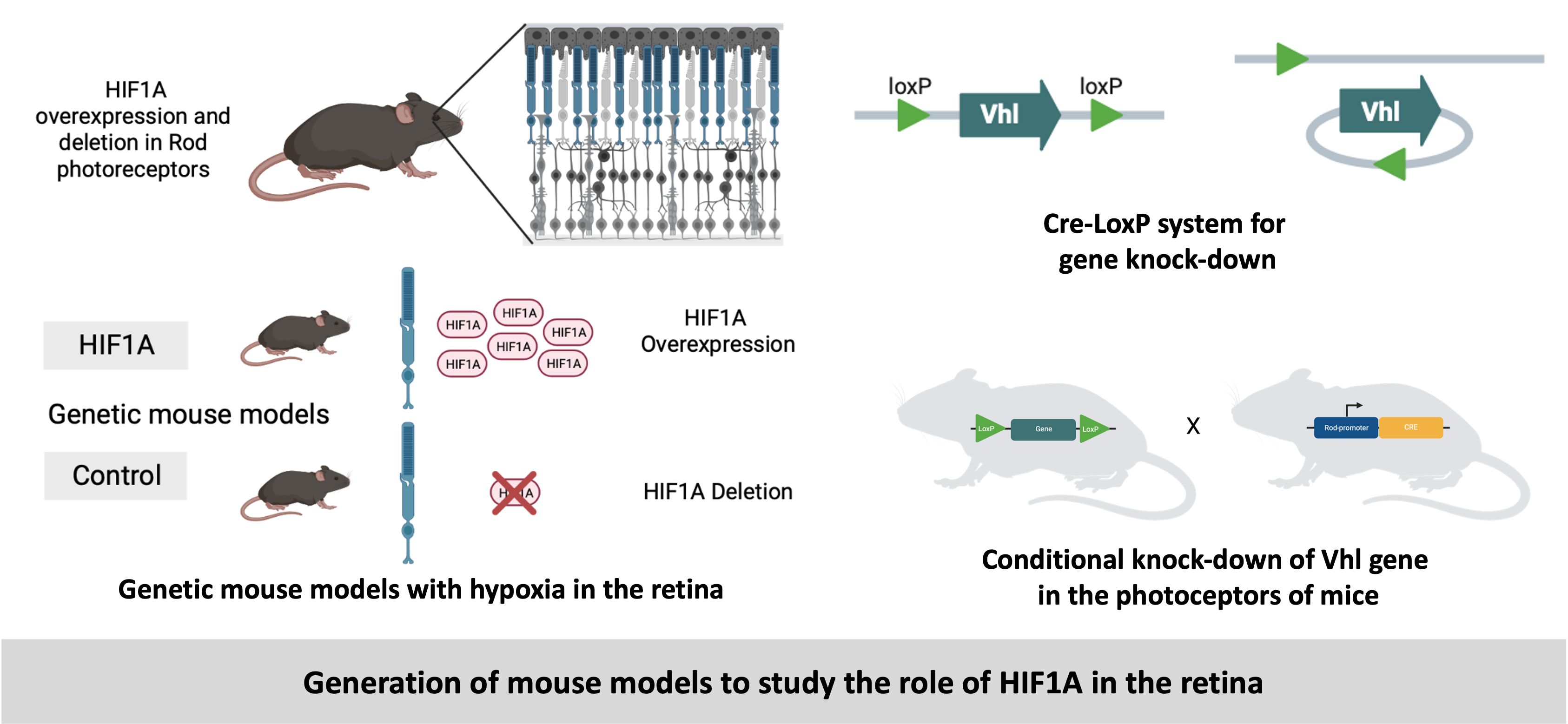
-
Read the details of my projects using rodent models -
- Inducible non-human primate models of retinal degeneration for testing end stage therapies and understanding disease mechanisms
Ail D, Nava D, Hwang I, Brazhnikova E, Nouvel-Jaillard C, Dentel A, Joffris C, Rousseau L, Dégardin J, Bertin S, Sahel JA, Goureau O, Picaud S, Dalkara D
Science Advances, 2023. Access paper here - Hif1a inactivation rescues photoreceptor degeneration induced by chronic hypoxia-like stress
Barben M*, Ail D*, Storti F, Klee K, Schori C, Samardzjia M, Michalakis S, Biel M, Meneau I, Blaser F, Barthelmes D, Grimm C
Cell Death and Differentiation, 2018. Access paper here - Increased expression of proton-sending G protein-coupled receptor Gpr65 during retina degeneration
Ail D*, Rüfenacht V*, Caprara C, Samardzjia M, Grimm C
Neuroscience, 2015. Access paper here - Linking YAP to Muller glia quiescence exit in the degenerative retina
Hamon A*, Ail D*, Garcia D*, Bitard J, Chesneau A, Dalkara D, Locker M, Roger J, Perron M
Cell Reports, 2019. Access paper here - Stanniocalcin2, but not Stanniocalcin1, responds to hypoxia in a HIF1-dependent manner in the retina
Ail D, Samardzija M, Chang ACM, Keck J, Reddel RR., Grimm C
Frontiers in Neuroscience, 2022. Access paper here - P38 MAPK signaling acts upstream of LIF-dependent neuroprotection during photoreceptor degeneration
Agca C, Gubler A, Traber G, Beck C, Imsand C, Ail D, Caprara C, Grimm C
Cell Death and Disease, 2013. Access paper here
Amphibians (Frog)
Frogs, particularly species like Xenopus laevis and Xenopus tropicalis, have served as valuable animal models in developmental biology due to their external development, large embryos, and ease of genetic manipulation. Their embryos are transparent and develop rapidly, allowing scientists to observe developmental processes in real time. Beyond development, Xenopus are widely being used as models for studying regeneration due to their remarkable ability to regrow damaged or lost tissues, especially in early life stages. Tadpoles can regenerate complex structures such as the tail, limbs, lens, spinal cord, and parts of the heart and brain and adult frogs also retain these regenerative capacities but to a limited extent. This ability makes them ideal models for uncovering the molecular and cellular mechanisms that drive regenerative processes.
We used these frogs as models to study retinal regeneration due to their natural ability to regenerate
retinal tissue after injury. Unlike mammals (like mice and humans), frogs can regenerate entire retinal layers, including photoreceptors and retinal ganglion cells, following damage or surgical removal. Our goal was to understand how and why are frogs able to regenerate their retina while humans cannot and what are the molecular mechanisms involved in this ability. The specific focus of this study was the Hedgehog signaling pathway and its key transcription factor called YAP.
We developed different inducible models to cause injury and degeneration to the retina. After injury/ degeneration was induced the process of regenration was followed and the molecular mechnisms involved were deciphered. A simple needle poke stratergy was used to cause a local injury in the retinas of both tadpoles and adult frogs. This technique allowed us to follow the degeneration and regeneration mechanisms in a small confined region of the retina (local pertubations and responses).
However, in order to analyze responses in a frog model more closely resembling human pathologies such as retinitis pigmentosa, we next generated a transgenic line in which rod photoreceptors can be specifically and conditionally ablated. We chose the Nitroreductase/Metronidazole (Ntr/Mtz) system previously used in Xenopus and modified it by fusing GFP to the NTR, to report transgene expression. The GFP allowed us to verify the expression of the NTR gene and when these tadpoles and frogs were subjected to treatment with the chemical metronidazole (MTZ), it induced loss of the photoreceptors. This technique allowed us to follow degeneration and regeneration mechanisms in the entire retina (global pertubations and responses).
Further, we created other models that deleted or overexpressed the transcrition factor YAP to specifically test the impact of YAP on the regeneratio mechanism. This was achieved by using a transgeneic line of Xenopus that expressed a dominant negative version of YAP. This was designed to be induced upon heta-shock treatment. This line was crossed with the NTR-MTZ line. The resulting offsprings could be chemically treated (MTZ) to cause degeneration and treatted with heat-shock for YAP deletion. To create transgenic Xenopus that overexpresed YAP we used a YAPS98A construct. The Ser- 98 residue, which is the target of Lats phosphorylation (Ser-127 in mammalian YAP), has been substituted with alanine (S98A). YAPS98A is therefore insensitive to Hippo pathway-mediated inhibition and thus constitutively active. This YAPS98A construct is fused to the ligand-binding domain of the glucocorticoid receptor (GR), which allows its conditional activation upon addition of dexamethasone (Dex).
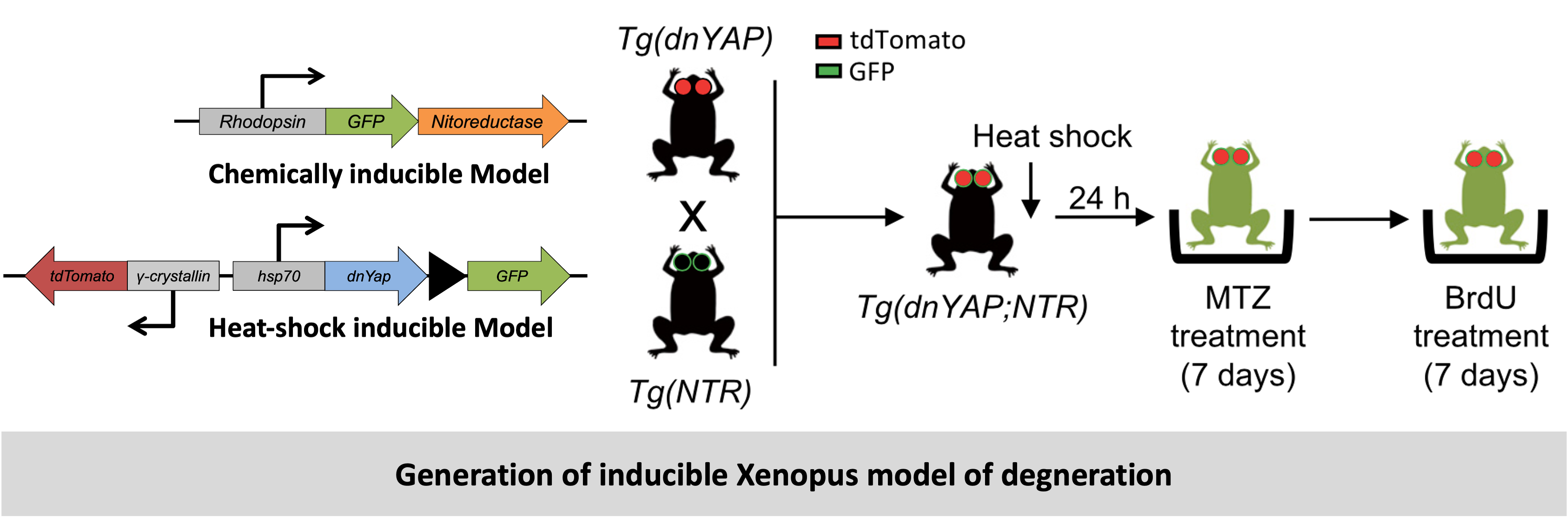
-
Read the details of my projects using frog models -
- Retinal degeneration and regeneration – Lessons from fishes and amphibians
Ail D and Perron M
Current Pathobiology Reports, 2017. Access paper here - Müller glial cell reactivation in Xenopus models of retinal degeneration
Langhe R*, Chesneau A*, Colozza G, Hidalgo M, Ail D, Locker M, Perron M
Glia, 2016. Access paper here - Linking YAP to Muller glia quiescence exit in the degenerative retina
Hamon A*, Ail D*, Garcia D*, Bitard J, Chesneau A, Dalkara D, Locker M, Roger J, Perron M
Cell Reports, 2019. Access paper here